Introduction
EPSP-to-Spike (E-S) potentiation has been observed in the rat hippocampus following stimulation trains which lead to LTP in the CA1 and dentate granule cell layer. Most commonly, E-S potentiation is noted as a potentiation in the population spike amplitude in excess of that predicted by the potentiation of the field EPSP slope. Jester, et al. (J Physiol, 1995) reported an associative stimulation paradigm which directly produces an ES potentiation with no change to the EPSP slope. Further work with field potential recordings and GABAB antagonists suggest that GABAB receptors are necessary for the expression of this associative E-S potentiation. What are the intracellular correlates of associative E-S potentiation and in what way are GABAB receptors involved in its expression?
Intracellular recordings were made from 75 g male Sprague-Dawley rats using sharp electrodes filled with 2M potassium acetate. In order to measure Na spike excitability and accommodation, a series of intracellular depolarizing 150 msec D-C current injections were given at approximately 5 minute intervals. The population spike amplitude and EPSP slope were monitored every 20 seconds with a single shock to the Schaffer-collateral pathway . The associative tetanization consisted of 50 bursts of 5 antidromic pulses at 100 Hz with an interburst interval of 200 ms, paired with one shock to the Schaffer-collaterals per antidromic burst.
In some cells, paired stimulation resulted in a slowly developing depolarization of the resting membrane potential (RMP) and an increase in spikes generated in the intracellular current injection series. Other cells seemed unaffected by the tetanization. Thus, it was not possible to detect a statistically significant effect of paired stimulation in RMP or input resistance in these recordings. Similarly, measurements of the GABAB mediated late IPSP were not affected by paired stimulation.
Methods
Standard methods were used to prepare hippocampal slices from 75g male Sprague-Dawley rats. Stimulating electrodes were placed on the Schaffer-collaterals to supply orthodromic activation and on the alveus to supply antidromic activation of the CA1 cell layer. The amplitude of the population spike and the peak initial slope of the EPSP were monitored by supplying orthodromic test shocks every 10 seconds.
The stimulus pattern used to generate associative E-S potentiation is shown on the top-left of the figure below. Fifty bursts of 5 pulses were delivered to the alveus with a 10 msec interpulse interval and a 200 msec interburst interval. A single orthodromic pulse is paired with the middle antidromic burst.
Intracellular potentials were amplified by an Axoclamp 2A amplifier (Axon Instruments) in bridge mode. Input resistance measures were taken from a 0.2 nA hyperpolarizing current injection. The GABAB-mediated late IPSP was measured as the difference in the membrane potential just before and 190 msec following a synaptic test shock.
Stimulation paradigm
Resting Membrane Potential
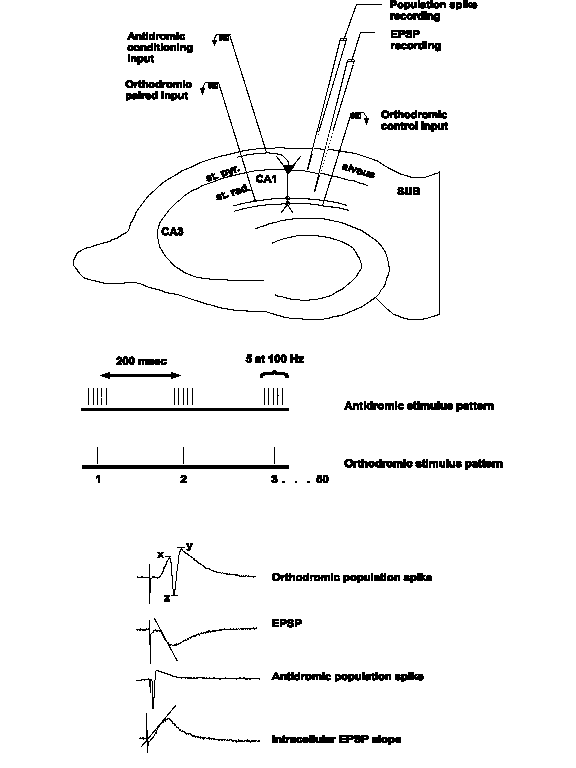
In this example, pairing anti- and orthodromic stimulation produced an E-S potentiation, seen as a population spike potentiation with no change in the slope of the EPSP or resting membrane potential (RMP).
Intracellular excitability increase
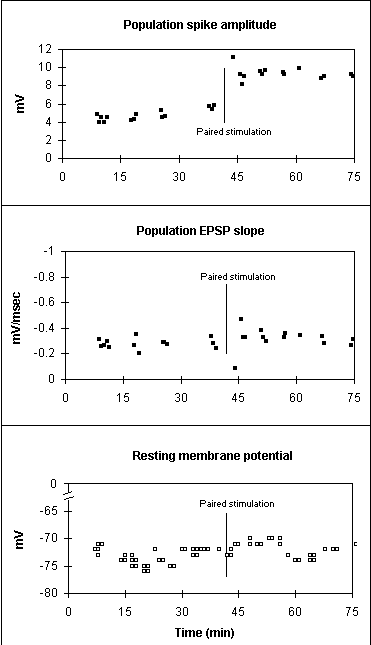
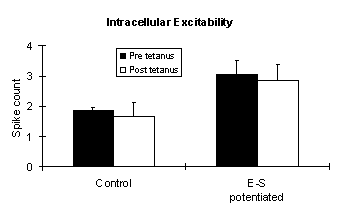
Paired stimulation has been shown to produce a slowly developing increase in intracellular excitability. The number of spikes counted during a series of depolarizing current injections, increased 15 min. after paired stimulation (Soc Neurosci Abstr 19:547.15) . Below are new results from 9 control and 4 ES potentiated slices showing the significant interaction between E-S potentiation and Pre vs. Post tetanus spike counts.
Below are results of a between/within split-plot analysis of variance (ANOVA). The between factor is control vs. E-S potentiated, and the within factor is pre vs. post tetanus. The control group is a combination of slices that only received antidromic tetanization plus slices that received paired stimulation but failed to exhibit ES potentiation.
Source Deg. Freedom F P Within subjects 13 Pre/Post tetanus 1 3.068 0.1077 E-S/(Pre/Post) 1 6.848 0.0241 Interaction
Input resistance
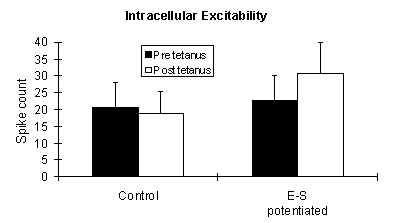
E-S potentiation had no effect on pre vs. post tetanus input resistance measurements. The means ± S.E.M are above and the results of a split-plot ANOVA are below for 4 control and 2 E-S potentiated slices. The between factor is control vs. E-S potentiated and the within factor is pre vs. post tetanus.
Source Deg. Freedom F P Within subjects 6 Pre/Post tetanus 1 0.912 0.3938 E-S/(Pre/Post) 1 0.007 0.9380 Interaction
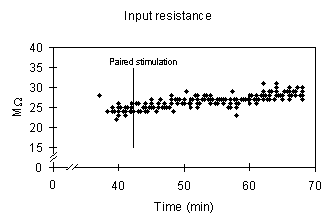
Input resistance, in this example, is not affected by a paired stimulation that resulted in an E-S potentiation of the population spike amplitude.
Intracellular EPSP slope in cells exhibiting E-S potentiation is also not affected by the E-S potentiation.
Tetanus N Intracellular EPSP slope Std. Error PRE 5 3.606 0.369 POST 5 3.670 0.416
GABAB block
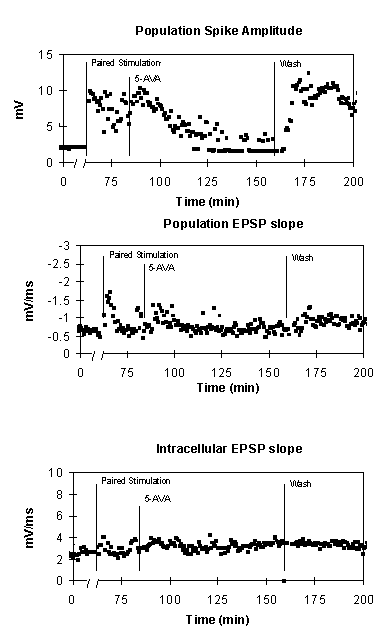
The GABAB blocker 5-aminovaleric acid (5AVA) blocks the expression of E-S potentiation. The top graph shows the potentiation of the population spike amplitude following paired stimulation, the reversal of that potentiation by 5AVA, and restoration of the potentiation upon 5AVA wash out. The middle and lower graphs show that the extracellular EPSP slope and intracellular EPSP slope are largely unaffected by either tetanization or medium change.
Late IPSP
To test whether the late IPSP, which is mediated by GABAB, was affected by E-S potentiation, the amplitude of the IPSP at 190 msec was compared before and after tetanization in both control and E-S potentiated slices.
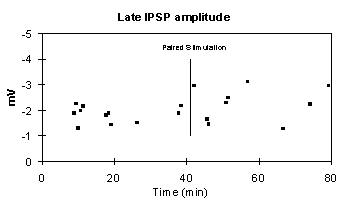
Late IPSP amplitude
Source Deg. Freedom F P
Within subjects
Pre/Post tetanus 1 0.340 0.578
1 0.000 0.997
E-S/(Pre/Post)Interacti
on
The results of a split-plot ANOVA in 4 control and 5 E-S potentiated slices show no difference between pre vs. post tetanus IPSPs and no interaction with E-S potentiation. Between subjects results are not shown. The example below shows the time-course of IPSP amplitude following E-S potentiation.
Recurrent inhibition, including the late IPSP, could possibly influence the intracellular excitability experiments since the number of spikes generated during a depolarization is somewhat dependent on spike frequency accommodation. These results suggest, however, that changes in the amplitude of the late IPSP did not influence intracellular excitability.
1. E-S potentiation is accompanied by an increase in intracellular excitability in response to depolarizing current injections.
3. The slopes of intracellular EPSPs were not affected by paired stimulation even in cases where intracellular excitability and input resistance were increased.
4. The GABAB blocker 5-aminovaleric acid blocked the expression of E-S potentiation. The GABAB-mediated late-IPSP was not significantly affected by ES potentiation.