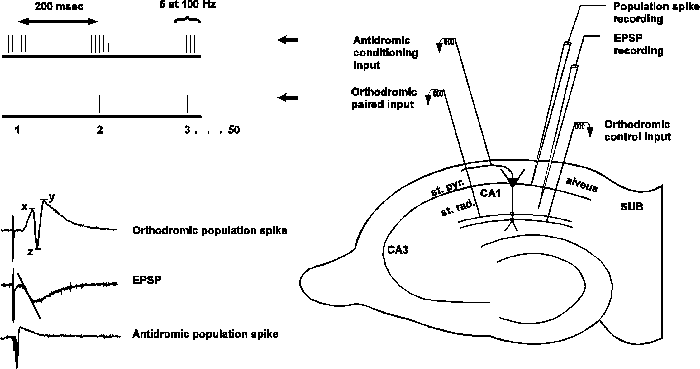
EPSP-to-spike or E-S potentiation is a component of LTP which results from a shift in the excitation /inhibition balance. In the presence of bicuculline, a GABAA antagonist, most of the E-S potentiation component of LTP is eliminated. Pairing antidromic theta-burst stimulation in phase with 5 Hz orthodromic stimulation produces an E S potentiation without affecting EPSP slope (Soc Neurosci Abstr 17:533.15). This associative form of E-S potentiation is not blocked by GABAA antagonists and is mechanistically distinct from the E-S potentiation of LTP (Soc Neurosci Abstr 19:547.15). Is GABAB involved in the induction or expression of antidromically-conditioned associative E-S potentiation? The EPSP and population spike were recorded from the CA1 layer of rat hippocampal slices. The antidromic conditioning stimulus was 50 bursts of 5 pulses at 100 Hz with an interburst interval of 200 ms delivered to the alveus. The Schaffer collaterals were stimulated at 5 Hz. The compound 5-aminovaleric acid (5AVA) was used in the bath to block GABAB receptors. Several results suggest GABAB receptors are involved in the expression, but not the induction, of associative E-S potentiation. 1) In slices showing E-S potentiation following paired stimulation, adding 1 mM 5AVA 10 min after pairing reduced the potentiation of the population spike to baseline; washing in normal bathing medium restored the potentiation to its former value. 2) In slices failing to potentiate following paired stimulation and in slices given the antidromic conditioning separate from the orthodromic stimulation, 5AVA had no effect on the population spike. 3) Delivering paired stimulation in 5AVA medium resulted in no E-S potentiation; washing out 5AVA 10 min after pairing unmasked the potentiation.
Standard methods were used to prepare hippocampal slices from 75g male Sprague-Dawley rats. Stimulating electrodes were placed on the Schaffer-collaterals to supply orthodromic activation and on the alveus to supply antidromic activation of the CA1 cell layer. The amplitude of the population spike and the peak initial slope of the EPSP were monitored by supplying orthodromic test shocks every 10 seconds.
The stimulus pattern used to generate associative E-S potentiation is shown on the top left of the figure below. Fifty bursts of 5 pulses are delivered to the alveus with a 10 msec interpulse interval and a 200 msec interburst interval. A single orthodromic pulse is paired with the middle antidromic burst. In some experiments, the orthodromic and antidromic stimulus trains were given separately to control for unpaired effects.
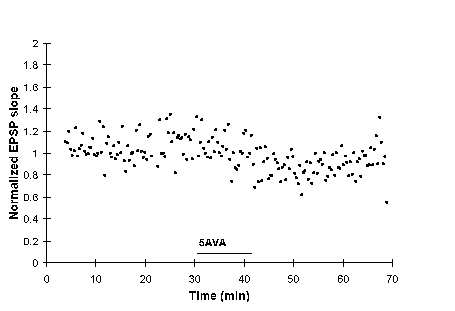
Associative E-S potentiation was generated by delivering paired orthodromic and antidromic stimulation (P). The decline in the baseline population spike amplitude (PSA) is not in response to tetanization: test shocks were delivered at 0.1 Hz. At 31 min, slices were perfused with medium containing 1 mM 5AVA, resulting in a return to baseline of PSA. Upon washout of 5AVA at 41 minutes, POP approached its former potentiated value. The GABAB blocker 5AVA reversibly blocked the expression of associative E-S potentiation. The bar drawn in the interior of the graph indicates the presence of 5AVA in the bathing medium.
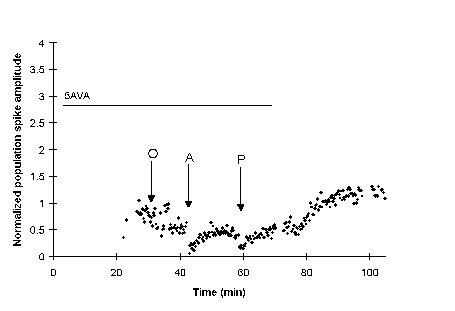
The antidromic and orthodromic stimulus trains, applied separately (A, O) or paired together (P), produce no lasting effect on the population spike of a slice bathed in 1 mM 5AVA until the 5AVA is washed out. This suggests slices exposed to 5AVA may be potentiated following a paired stimulus, but that the expression of the potentiation is masked.
To control for the possibility that 5AVA has an effect on the baseline population spike amplitude, rather than the expression of associative E-S potentiation, slices that did not receive paired stimulation were exposed to 5AVA. The orthodromic and antidromic stimuli were delivered separately (A, O) during exposure to 5AVA. Since in this control, there is no rebound upon washout of 5AVA, the rebound seen in panel 4 is likely a consequence of the paired stimulation rather than just the antidromic or orthodromic stimulation alone. The only potentiation seen in either trace in this panel is a temporary potentiation in response to the orthodromic stimulation, and a slight depression following the antidromic stimulation.
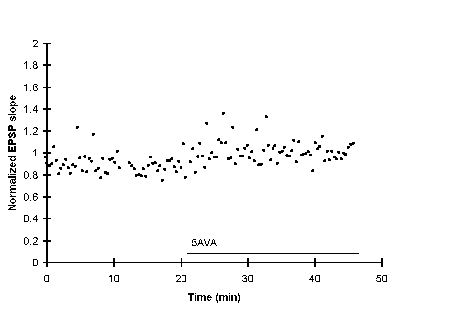
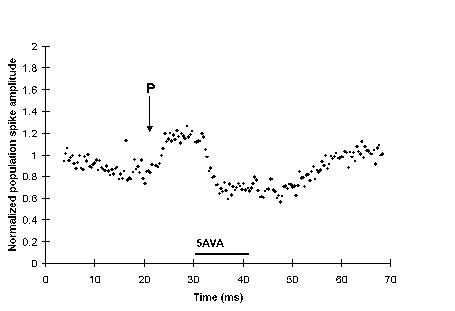
A paired stimulation (P) was given at 16 min, but E-S potentiation failed to develop. Perfusing with medium containing 5AVA following this failed attempt to induce E-S potentiation had almost no effect on the population spike amplitude suggesting that 5AVA reduces the population spike amplitude only when it has been potentiated by a successful paired stimulation.
Since data in this poster suggest that GABAB receptors are involved in the expression of associative E-S potentiation, it is likely that associative E-S potentiation and paired pulse stimulation will interact, since GABAB receptors are known to contribute to paired pulse facilitation.